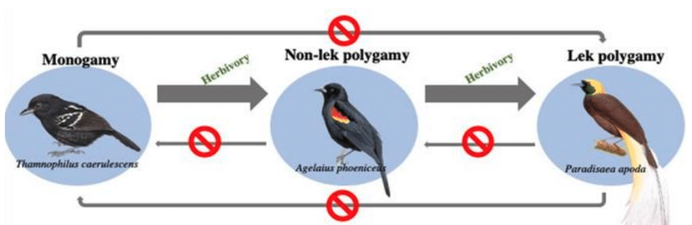
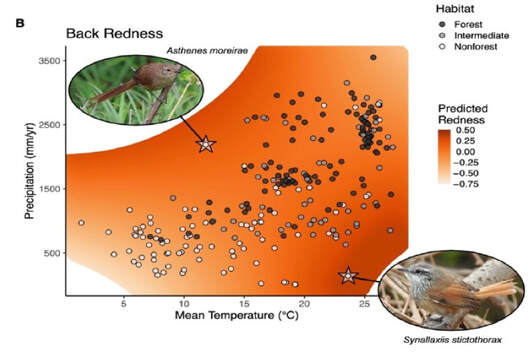
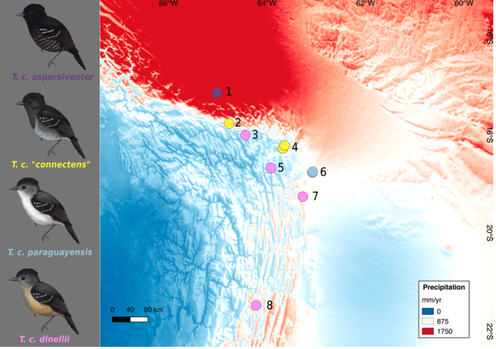
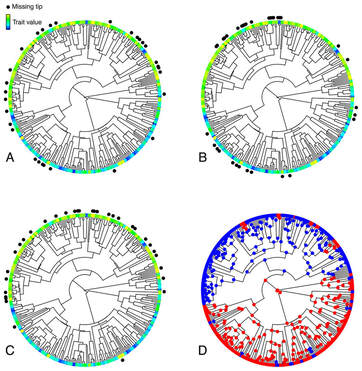
For scientists and laypersons alike, birds have always held a special place in the pantheon of animal forms. Their ubiquity, mostly diurnal habits, often vivid colors, and melodious vocalizations make them particularly conspicuous and charismatic to human sensibilities. Accordingly, they have been at the center of many of the most important conceptual advances in evolutionary biology. My research goals follow on that tradition, and center around understanding the processes that have given rise to the phenotypic diversity of birds.*
Below I describe a few of the specific topics I'm currently working or have recently worked on. For a complete and up-to-date list of my publications, check out my Google Scholar profile. Please do not hesitate to contact me if you would like a pdf of one of my works.
*The preceding lines are adapted from RS Marcondes. 2022. Celebrating the great avian evolutionary tradition: Review of how birds evolve, by DJ Futuyma, Princeton & Oxford: Princeton University Press. Evolution 76: 1366-1369.
Below I describe a few of the specific topics I'm currently working or have recently worked on. For a complete and up-to-date list of my publications, check out my Google Scholar profile. Please do not hesitate to contact me if you would like a pdf of one of my works.
*The preceding lines are adapted from RS Marcondes. 2022. Celebrating the great avian evolutionary tradition: Review of how birds evolve, by DJ Futuyma, Princeton & Oxford: Princeton University Press. Evolution 76: 1366-1369.